Raaf
Die raaf (Corvus corax) is ’n groot, swart sangvoël. Hulle word in die Noordelike Halfrond aangetref en is die lede van die familie Corvidae wat die wydste verspreid is. Daar is minstens agt subspesies wat nie veel in voorkoms verskil nie, maar daar is aansienlike genetiese verskille tussen die bevolkings van verskillende streke. Dit is een van die grootste Corvidae-spesies en moontlik die swaarste sangvoël; ’n volwasse raaf is sowat 63 cm lank en weeg omtrent 1,2 kg.
| Raaf | |
|---|---|

| |
| C. corax tibetanus afgeneem in Tibet en 'n klankopname uit Italië | |
| Wetenskaplike klassifikasie | |
| Koninkryk: | |
| Filum: | |
| Klas: | |
| Orde: | |
| Familie: | |
| Genus: | |
| Spesie: | C. corax (Linnaeus, 1758)
|
| Binomiale naam | |
| Corvus corax | |

| |
| verspreiding | |
Hulle kan 21 jaar lank in die natuur oorleef.[1] Jong voëls kan in swerms voorkom, maar hulle vorm daarna pare vir die res van hul lewe.
Rawe bestaan al duisende jare lank saam met die mens en is in sommige gebiede so talryk dat hulle as ’n plaag beskou word. Deel van hul sukses as ’n spesie is vanweë hul omnivore dieet. Hulle is ook baie goed daarin om kos te vind, veral aas, insekte, graan, bessies, vrugte, klein diertjies en afvalkos.
Rawe is buitengewoon intelligent.[2] Hulle was deur die eeue die onderwerp van mitologie, folklore, kuns en letterkunde. In baie kulture, soos die inheemse kulture van Skandinawië, Ierland, Wallis, Bhoetan, Noord-Amerika en Siberië, is die raaf beskou as ’n spirituele figuur of godheid.[3]
Evolusie
wysigDie raaf het in die Ou Wêreld ontstaan en oor die Beringlandbrug na Noord-Amerika migreer.[4] Onlangse genetiese studies, waarin die DNS van rawe oor die wêreld heen ondersoek is, het daarop gedui rawe val in minstens twee klades: ’n Kaliforniese klade, wat in die suidweste van die VSA aangetref word, en ’n Holarktiese klade, wat in die res van die Noordelike Halfrond voorkom. Die voëls van die twee klades lyk eenders, maar is geneties verskillend; hulle het sowat 2 miljoen jaar gelede van mekaar begin skei.[5][6]
Daar is in toetse met mitochondriale DNS bevind die rawe van die res van Amerika is nader verwant aan dié van Eurasië as aan dié in die Kaliforniese klade, en dat die rawe van die Kaliforniese klade nader verwant is aan die woestynraaf (C. cryptoleucus) as aan dié van die Holarktiese klade.[5] Rawe in die Holarktiese klade is nader verwant aan die witborskraai (C. albus) as aan die Kaliforniese klade.[7]
’n Verduideliking vir hierdie genetiese bevindings is dat rawe hulle minstens 2 miljoen jaar gelede in Kalifornië gevestig het en tydens ’n ystydperk geskei geraak het van hul verwante in Eurasië. ’n Miljoen jaar gelede het ’n groep van die Kaliforniese klade in ’n nuwe spesie, die woestynraaf, begin ontwikkel. Ander lede van die Holarktiese klade het later in ’n aparte migrasie uit Asië gearriveer, miskien saam met die mens.[8]
’n Onlangse studie met mitochondriale DNS het getoon die geïsoleerde bevolking van die Kanariese Eilande verskil van ander bevolkings.[9] Die studie het nie voëls van Noord-Afrika ingesluit nie[9] en hul posisie is dus nie duidelik nie, maar hul morfologie stem baie ooreen met die rawe van die Kanariese Eilande (in so ’n mate dat die twee dikwels as een enkele subspesie beskou word).[10]
Klassifikasie
wysigDie naaste verwant aan die raaf is die bruinnekraaf (C. ruficollis), witborskraai (C. albus) van Afrika en woestynraaf (C. cryptoleucus) van die suidweste van Noord-Amerika.[11] Sommige owerhede erken tot 11 subspesies,[12] maar ander erken net agt:[10]
| Subspesie | Foto | Verspreiding | Notas |
|---|---|---|---|
| C. c. corax | 
|
Van Europa ooswaarts tot die Baikalmeer, suid tot die Kaukasus en Noord-Iran | Dit het ’n relatief kort, geboë snawel. Die bevolkings van Suidwes-Europa het ’n selfs geboër snawel en korter vlerke as gemiddeld, en daarom erken sommige owerhede dit as ’n aparte subspesie, C. c. hispanus.[12] |
| C. c. varius | 
|
Ysland en die Faroëreilande | Dit is minder blink as C. c. principalis of corax, is van gemiddelde grootte en die basis van sy nekvere is witterig (nie op ’n afstand sigbaar nie). |
| C. c. subcorax | 
|
Van Griekeland ooswaarts tot by Noordwes-Indië, Sentraal-Asië en Wes-China, maar nie in die Himalaja nie | Dit is groter as gemiddeld, maar het relatief kort nekvere. Dit is hoofsaaklik swart, hoewel sy nek en bors nes die bruinnekraaf s’n ’n bruinerige tint het. Die basis van sy nekvere is dikwels amper witterig, hoewel die kleur kan wissel. |
| C. c. tingitanus | 
|
Noord-Afrika en die Kanariese Eilande | Dit is die kleinste subspesie, met die kortste nekvere en ’n olierige veerglans. Sy snawel is kort, maar stewig en geboë. Dié van die Kanariese Eilande is bruiner as dié van Noord-Afrika en daarom beskou sommige wetenskaplikes hulle as aparte subspesies, onderskeidelik C. c. canariensis en C. c. tingitanus.[12] |
| C. c. tibetanus | 
|
Himalaja | Dit is die grootste en blinkste subspesie, met die langste nekvere. Sy snawel is groot, maar minder indrukwekkend as dié van C. c. principalis, en die basis van sy nekvere is grys. |
| C. c. kamtschaticus | 
|
Noordoos-Asië | Dit is van gemiddelde grootte, tussen C. c. principalis en C. c. corax, en het ’n dikker en groter snawel as gemiddeld. |
| C. c. principalis | 
|
Noordelike Noord-Amerika en Groenland | Dit het ’n groot lyf en die grootste snawel. Sy vere blink baie en sy nekvare is goed ontwikkel. |
| C. c. sinuatus | 
|
Suidelike Sentraal-VSA en Sentraal-Amerika. | Dit is kleiner, met ’n kleiner en smaller snawel as C. c. principalis. Bevolkings in die verre suidwestelike VSA en Noordwes-Mexiko is die kleinstes in Noord-Amerika. Hulle word soms ingesluit in C. c. sinuatus, terwyl ander owerhede hulle beskou as ’n aparte subspesie, C. c. clarionensis.[12] |
Beskrywing
wysig
’n Volwasse raaf is tussen 54 en 67 cm lank, met ’n vlerkspan van 115 tot 130 cm.[13][14][15][16] Die gewig wissel van 0,69 tot 2 kg[17][18] en daarom is die raaf een van die swaarste sangvoëls. Gewig tussen subspesies verskil: Rawe van Kalifornië weeg gemiddeld 784 g, dié van Alaska gemiddeld 1,13 kg en dié van Nova Scotia 1,23 kg.[19][20][21] Die snawel is groot en effens geboë, met ’n lengte van 5,7 tot 8,5 cm; dit is van die grootste snawels onder die sangvoëls.
Rawe het ’n langerige stert, sowat 20 tot 26,3 cm, met hoofsaaklik swart vere. Die nekvere is lank en gepunt en die basis daarvan is ’n vaal bruingrys. Die pote is ’n redelike lengte, met ’n voetwortel van 6 tot 7,2 cm lank.[22][23]
Benewens dat hy groter is, verskil die raaf van sy neef die kraai vanweë sy groter en swaarder swart snawel, wollerige vere om die keel en bo die snawel en ’n wigvormige stert.[24] Vlieënde rawe kan van kraaie onderskei word aan hul stertvorm, groter vlerke en stabieler opstygstyl, met minder van ’n vlerkgeklap. Rawe se stem is ook eiesoortig – sy roep is ’n diep gekras wat baie helderder is as dié van ’n kraai. Altesaam 15 tot 30 verskillende geluide is al aangeteken, waarvan die meeste vir sosiale interaksie gebruik word.[25] Nes ander Kraaiagtiges kan rawe klanke uit die omgewing namaak, insluitende die stem van mense. Nievokale klanke sluit in fluitgeluide met die vlerke en klapgeluide met die snawel. As ’n raaf weg is, sal sy maat sy roep namaak om hom aan te spoor om terug te keer.[26]
Verspreiding en habitat
wysig
.jpg)

Rawe kan in enige klimaat floreer; hierdie spesie het inderdaad die grootste verspreiding van al die lede van die genus[27][28] en een van die grootstes van alle sangvoëls.[29] Hulle kom voor dwarsdeur die Noordelike Halfrond, van arktiese tot gematigde habitats in Noord-Amerika en Eurasië tot die woestyne van Noord-Afrika, en die eilande in die Stille Oseaan.
Die meeste verkies beboste gebiede met groot, oop velde daarnaby of kusstreke vanweë hul plekke om nes te maak en kos te kry. In sommige digbewoonde gebiede soos Kalifornië slaan hulle munt uit die algemene beskikbaarheid van kos en hul getalle neem daar toe.[30] Aan die kus is hulle meer eweredig versprei en bou hulle hul neste teen kranse.[31] Rawe kom dikwels aan die kus voor, want kos en water is geredelik beskikbaar.[31] Die weer en temperatuur is ook stabiel.
Gedrag
wysigIntelligensie
wysigDie raaf se brein is van die grootste van alle voëls. Hulle toon ’n vermoë om probleme op te los, sowel as ander kognitiewe prosesse (om te leer en kennis op te doen) soos nabootsing en insig.[32]
Die linguis Derek Bickerton, wat voortgebou het op werk deur die bioloog Bernd Heinrich, het gereken rawe is een van net vier bekende diere (die ander drie is bye, miere en mense) wat kan kommunikeer oor voorwerpe of gebeure wat ver in tyd of ruimte verplaas is (sg. linguistiese verplasing). Jong rawe sonder paarmaats sal snags saam slaap, maar bedags alleen na kos gaan soek. As een van hulle op ’n karkas afkom wat deur ’n volwasse paar rawe opgepas word, sal die jong raaf dié aand na die ander terugkeer en vir hulle van sy vonds vertel. ’n Groep jong rawe sal dan die volgende dag na die karkas vlieg en die volwasse paar wegjaag. Volgens Bickerton was die ontwikkeling van linguistiese verplasing dalk die belangrikste gebeure in die evolusie van die menslike taal en is rawe en mense die enigste gewerweldes met dié vermoë.[33]
Een eksperiment wat ontwerp is om insig en probleemoplossing te toets, was om ’n stuk vleis aan ’n tou vas te maak wat van ’n dwarsstok afhang. Om die vleis in die hande te kry moes die raaf op die dwarsstok staan, die tou entjie vir entjie optrek en op die lusse trap om die tou geleidelik korter te maak. Vier of vyf rawe het eindelik daarin geslaag, en die vordering van geen sukses (deur net aan die tou te pluk) tot sukses (deur die vleis te kry) het plaasgevind sonder enige teken van ’n leer-en-probeer-proses. Dit steun die hipotese dat rawe "uitvinders" is wat probleme kan oplos. Voorheen is gereken hul gedrag is aangebore, maar daar is nou vasgestel hul vermoë om probleme individueel op te los en van mekaar te leer, weerspieël ’n buigsame kapasiteit vir intelligente insig wat ongewoon is onder diere.[34]
’n Ander eksperiment dui daarop dat rawe ten minste ander rawe met opset kan flous.[35]
Hulle kyk waar ander rawe hul kos bêre, onthou die bêreplek en kan later die kos gaan steel. Dit gebeur so dikwels dat rawe soms ver weg van die kosplek sal vlieg om ’n beter wegsteekplek te kry.[36] Daar is ook al waargeneem dat hulle net maak of hulle hul kos op ’n sekere plek wegsteek om ander rawe om die bos te lei.[37]
Rawe is bekend daarvoor dat hulle blink voorwerpe soos skulpe, stukkies metaal en golfballe steel en wegsteek. Een teorie is dat hulle blink voorwerpe bymekaarmaak om ander rawe te beïndruk.[38] In ’n ander studie dui die bevindings daarop dat jong rawe uiters nuuskierig is oor alle nuwe dinge en dat hulle van blink, ronde voorwerpe hou vanweë hul ooreenkoms met voëleiers. Volwasse rawe verloor dié belangstelling in die ongewone en ontwikkel ’n groot vrees vir alles wat nuut is.[39]
Spel
wysigDie belangstelling neem toe in die mate waarin voëls speel. Jong rawe is van die spelerigstes van alle voëlsoorte. Hulle is al gesien waar hulle teen sneeubanke afgly, blykbaar net vir die pret. Hulle speel selfs speletjies met ander diere, soos vang-my-as-jy-kan met wolwe, otters en honde.[40] Rawe is bekend vir hul skouspelagtige aërobiese vertonings, soos om saam in lusse of met ineengehaakte kloue te vlieg.[41][42]
Hulle is ook een van die min wilde diere wat hul eie speelgoed maak. Daar is al gesien hoe hulle takkies afbreek om gesellig mee te speel.[43]
Verwysings
wysig- ↑ Wasser, D. E.; Sherman, P.W. (2010). "Avian longevities and their interpretation under evolutionary theories of senescence". Journal of Zoology. 280 (2): 103–155. doi:10.1111/j.1469-7998.2009.00671.x.
- ↑ http://www.abc.es/ciencia/abci-cuervos-pueden-inteligentes-como-nino-tres-anos-201604262232_noticia.html
- ↑ Jones, Noragh (1995). Power of Raven, Wisdom of Serpent. Floris Books. ISBN 0-940262-66-5.
- ↑ Marzluff and Angell, p. 86
- ↑ 5,0 5,1 US Geological Survey. "California Ravens Are a Breed Apart" (in Engels). Geargiveer vanaf die oorspronklike op 8 Julie 2017. Besoek op 11 Mei 2007.
- ↑ Omland KE; Tarr CL; Boarman WI; Marzluff JM; Fleischer RC (2000). "Cryptic genetic variation and paraphyly in ravens". Proceedings of the Royal Society B. 267: 2475–82. doi:10.1098/rspb.2000.1308. PMC 1690844. PMID 11197122.
- ↑ Feldman, Christopher R.; Omland, Kevin E. (Maart 2005). "Phylogenetics of the common raven complex (Corvus: Corvidae) and the utility of ND4, COI and intron 7 of the β-fibrinogen gene in avian molecular systematics". Zoologica Scripta. 34 (2): 145–156. doi:10.1111/j.1463-6409.2005.00182.x.
- ↑ Marzluff and Angell, pp. 86–87
- ↑ 9,0 9,1 Baker, Jason M.; Omland, Kevin E. (Januarie 2006). "Canary Island Ravens Corvus corax tingitanus have distinct mtDNA". Ibis. 148 (1): 174–178. doi:10.1111/j.1474-919X.2006.00493.x.
- ↑ 10,0 10,1 Clements, J. F. (2007). The Clements Checklist of the Birds of the World. 6de uitg. Christopher Helm. ISBN 978-0-7136-8695-1
- ↑ Goodwin, pp. 70–72
- ↑ 12,0 12,1 12,2 12,3 Marzluff, J. M. (2009). "Common Raven (Corvus corax)". pp. 638–639 in Handbook of the Birds of the World. Bush-shrikes to Old World Sparrows. del Hoyo, J., Elliott, A. en Christie, D. A. (reds.). Lynx Edicions, Barcelona. ISBN 978-84-96553-50-7
- ↑ Svensson, Lars; Mullarney, Killian; Zetterström, Dan (2015). Le guide ornitho: le guide le plus complet des oiseaux d'Europe, d'Afrique du Nord et du Moyen-Orient [The birding guide: The most comprehensive guide of birds from Europe, North Africa and the Middle East]. Frans: Éditions Delachaux et Niestlé.
- ↑ Common Raven. Nature.ca. Besoek op 2012-12-19.
- ↑ Raven. Garden-birds.co.uk (2010-07-01). Afgelaai on 2012-12-19.
- ↑ Common Raven. Geobirds.com. Besoek op 2012-12-19.
- ↑ Boarman, William I.; Heinrich, Bernd (1999). Poole, A.; Gill, F. (reds.). "Common Raven (Corvus corax)". Birds of North America. 476: 1–32. doi:10.2173/bna.476.
- ↑ Common Raven Geargiveer 19 Desember 2017 op Wayback Machine. Oiseaux-birds.com. Besoek op 2012-12-19.
- ↑ Elliot, R. D. (1977). "Hanging behavior in Common Ravens". Auk. 94 (4): 777–778. doi:10.2307/4085278. JSTOR 4085278.
- ↑ Schwan, M. W.; Williams, D. D. (1978). "Temperature regulation in the common raven of interior Alaska". Comparative Biochemistry and Physiology A. 60: 31–36. doi:10.1016/0300-9629(78)90033-6.
- ↑ Linz, G. M., Knittle, C. E. en Johnson, R. E. (1990). Ecology of corvids in the vicinity of the Aliso Creek California Least Tern colony, Camp Pendelton, California. U.S. Dept. of Agric., North Dakota Field Stn. North Dakota State Univ., Fargo.
- ↑ Oberholser, Harry C. (1918). "The Common Ravens of North America". The Ohio Journal of Science. 18 (6): 213–225.
- ↑ Anonymous. (2013) "Corvus corax – Linnaeus, 1758 (Common Raven)" in Deomurari, A.N. (samesteller), 2010. AVIS-IBIS (Avian Information System – Indian BioDiversity Information System) v. 1.0. Foundation For Ecological Security, Indië
- ↑ Vere Benson, S. (1972). The Observer's Book of Birds. Londen: Frederick Warne & Co. Ltd. ISBN 0-7232-1513-8.
- ↑ Gwinner, E. (1964). "Untersuchungen über das ausdrucks und Sozialverhalten des Kolkraben (Corvus corax L.)". Zeitschrift für Tierpsychologie (in Duits). 21 (6): 657–748. doi:10.1111/j.1439-0310.1964.tb01212.x.
- ↑ Goodwin, p. 142
- ↑ Goodwin, p. 70
- ↑ Madge, Steve (1999) [1994]. Crows and jays : a guide to the crows, jays and magpies of the world. Helm Identification Guides. Londen: Christopher Helm. ISBN 0-7136-3999-7.
- ↑ "Common Raven". Audubon Society of Portland. 2012. Besoek op 6 November 2012.
- ↑ U.S. Geological Survey. "Scientists Estimate Risk of Raven Predation on Desert Tortoises in the Western Mojave Desert" (in Engels). Geargiveer vanaf die oorspronklike op 22 Desember 2015. Besoek op 11 Mei 2007.
- ↑ 31,0 31,1 Ewins, P. J.; Dymond, J. N.; Marquiss, M. (1986). "The distribution, breeding and diet of Ravens Corvus corax in Shetland". Bird Study. 33 (2): 110–116. doi:10.1080/00063658609476906.
- ↑ "PBS Nature: The Bird in Black" (in Engels). PBS. Geargiveer vanaf die oorspronklike op 17 Julie 2008. Besoek op 7 Mei 2007.
- ↑ Bickerton, Derek (2009). Adam's Tongue. Hill and Wang. ISBN 978-0-8090-2281-6.
- ↑ Heinrich, Bernd (1995). "An Experimental Investigation of Insight in Common Ravens (Corvus corax)" (PDF). The Auk. 112 (4): 994–1003. doi:10.2307/4089030. JSTOR 4089030. Besoek op 16 Mei 2007.
- ↑ Bugnyar, Thomas; Kotrschal, Kurt (2004). "Leading a conspecific away from food in ravens (Corvus corax)?" (PDF). Animal Cognition. 7 (2): 69–76. doi:10.1007/s10071-003-0189-4. Geargiveer vanaf die oorspronklike (PDF) op 28 September 2015.
- ↑ Rozell, Ned. "The Raven's Game of Hide and Seek". Alaska Science Forum (in Engels). Geophysical Institute, University of Alaska Fairbanks. Geargiveer vanaf die oorspronklike op 27 April 2012. Besoek op 7 Mei 2007.
- ↑ Marzluff and Angell, p. 230
- ↑ Marzluff and Angell, p. 232
- ↑ Kijne, M.; Kotrschal, K (2002). "Neophobia affects choice of food-item size in group-foraging common ravens (Corvus corax)". Acta ethologica. 5 (1): 13–18. doi:10.1007/s10211-002-0061-6.
- ↑ Savage, pp. 70–71
- ↑ Savage, p. 76
- ↑ Heinrich, B. (1999). Mind of the Raven: Investigations and Adventures with Wolf-Birds p. 290. New York: Cliff Street Books. ISBN 978-0-06-093063-9
- ↑ Heinrich, B. (1999). Mind of the Raven: Investigations and Adventures with Wolf-Birds p. 282. New York: Cliff Street Books. ISBN 978-0-06-093063-9
Bronne
wysig- Goodwin D. (1983). Crows of the World. Queensland University Press, St Lucia, Qld. ISBN 0-7022-1015-3.
- Marzluff, John M.; Angell, Tony (2005). In the Company of Crows and Ravens. New Haven: Yale Univ. Press. ISBN 0-300-10076-0.
- Savage, Candace (1995). Bird Brains: The Intelligence of Crows, Ravens, Magpies and Jays. Toronto: Douglas & McIntyre. ISBN 1-55054-189-7.
Verdere leesstof
wysig- Heinrich, B. (1999). Mind of the Raven: Investigations and Adventures with Wolf-Birds. New York: Cliff Street Books. ISBN 978-0-06-093063-9
Eksterne skakels
wysig- RSPB: Raaf
- Raaf-subspesies – Cornell Lab of Ornithology
- Foto's van rawe by ARKive Geargiveer 9 Mei 2007 op Wayback Machine
 Wikispecies het meer inligting verwant aan Raaf
Wikispecies het meer inligting verwant aan Raaf Wikimedia Commons het meer media in die kategorie Raaf.
Wikimedia Commons het meer media in die kategorie Raaf. Wikiwoordeboek het 'n inskrywing vir raaf.
Wikiwoordeboek het 'n inskrywing vir raaf.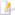 Hierdie artikel is vertaal uit die Engelse Wikipedia
Hierdie artikel is vertaal uit die Engelse Wikipedia