Koraal
Korale, of koraaldiertjies, is ongewerwelde seediertjies in die klas Anthozoa (blomdiere) van die filum Cnidaria. Hulle kom gewoonlik in kompakte kolonies van talle identiese, individuele poliepe voor. Die bekendste van die korale is die steenkorale (Scleractinia). Hulle het die hoofaandeel in die ontstaan van koraalriwwe, wat bestaan uit kalsiumkarbonaat wat die diertjies afskei om ’n harde skelet te vorm.
| Koraal | |
|---|---|
.jpg)
| |
| Steenkoraal. | |
| Wetenskaplike klassifikasie | |
| Koninkryk: | |
| Filum: | |
| Klas: | Anthozoa Ehrenberg, 1834
|
| Subklasse | |
Elke poliep in die kolonie is identies en bestaan uit ’n sakkie met ’n stel tentakels wat die sentrale opening omring. Hulle is gewoonlik ’n paar millimeter breed en ’n paar sentimeter lank. ’n Eksoskelet word naby die basis afgeskei. Oor baie geslagte heen skep die kolonie dus ’n groot skelet wat kenmerkend van die spesie is. Hulle kan aseksueel voortplant, of seksueel deur kuitskieting: poliepe van dieselfde spesie stel gelyktydig oor ’n tydperk van een tot verskeie nagte gamete vry, gewoonlik min of meer met volmaan.
Hoewel sommige korale klein vissies en plankton vang met die netelselle aan die punt van hul tentakels, kry die meeste hul energie en voedingstowwe van fotosintetiese, eensellige alge van die genus Symbiodinium bekend as soöxantellas wat in hul weefsel woon. Sulke korale het sonlig nodig en groei in helder, vlak water, gewoonlik op ’n diepte van minder as 60 meter. Korale dra baie by tot die fisiese struktuur van koraalriwwe wat in tropiese en subtropiese klimate ontwikkel, soos die Groot Koraalrif aan die kus van Queensland, Australië.
Ander korale steun nie op soöxantellas nie en kan in baie dieper water oorleef, met die kouewater-genus Lophelia wat tot op 3 000 meter kan voorkom.[4] Sommige is al noordwes van Cape Wrath in Skotland ontdek en ander so ver noord as aan die kus van die deelstaat Washington en die Aleoetiese Eilande.
Taksonomie en anatomie
wysig
| |||||||||||||||||||||||||||||||||||||||
| Afstamming van Anthozoa |
Korale word in die klas Anthozoa van die filum Cnidaria geklassifiseer. Hulle word in drie subklasse verdeel:
- Hexacorallia. Sluit die steenkorale, seeanemone en soantides in. Hierdie groepe het poliepe wat gewoonlik ses tentakels of ’n veelvoud van ses het.
- Octocorallia. Sluit bloukorale, sagte korale, seevere en horingkorale in. Hulle het agt tentakels elk.[5]
- Ceriantharia. Word ook viltkokeranemone genoem.[6][7][8]

Brandkorale is nie ware korale nie; hulle val in die orde Anthomedusa (soms bekend as Anthoathecata) van die klas Hydrozoa.[9]
Korale is steellose diertjies wat van die meeste ander lede van Cnidaria verskil deurdat hulle nie ’n kwalvormstadium in hul lewensiklus het nie. Die lyf van die diertjie is ’n poliep. Die meeste korale is deel van kolonies; een poliep produseer ’n ander een en die kolonie ontwikkel eindelik uit hierdie klein begin. By steenkorale produseer die poliep ’n skelet wat uit kalsiumkarbonaat bestaan om die organisme te versterk en beskerm. Dit word afgeskei deur die poliepe en die senosark, die lewende weefsel wat hulle verbind. Kolonies steenkorale kan baie verskil wat voorkoms betref; die verskillende vorms word dikwels verbind met verskillende soorte habitat, verskille in die hoeveelheid lig en die beweging van die water.[10]
Sagte korale het nie harde skelette nie, maar die weefsel word dikwels versterk deur die teenwoordigheid van klein skeletagtige elemente, skleriete, wat uit kalsiumkarbonaat bestaan. Sagte korale se vorms wissel baie en die meeste is koloniaal. ’n Paar sagte korale is stolonagtig, maar die poliepe van die meeste word verbind deur plate senosark. By sommige spesies is dit dik en die poliepe is heeltemal veranker. Sommige sagte korale is korsagtig of vorm lobbe. Ander is boom- of sweepagtig en het ’n sentrale aksiale skelet wat in die weefselbindmiddel veranker is.
In beide steen- en sagte korale kan die poliepe teruggetrek word, met steenkorale wat op hul harde skelet en netelselle staatmaak vir verdediging en sagte korale wat gewoonlik chemiese verdedigingsmetodes gebruik in die vorm van giftige stowwe wat in die weefsel aanwesig is.[10]

Die poliepe van steenkorale het ’n sesvoudige simmetrie en dié van sagte korale ’n agtvoudige simmetrie. Die mond van elke poliep word omring deur tentakels, of vangarms. By steenkorale is hulle silindries en loop dunner na die punt, maar by sagte korale is hulle getak met uitsteeksels bekend as pinnules. By sommige tropiese spesies is hulle net klein stompies en by ander saamgesmelt om soos ’n spaan te lyk.[11]
By die meeste korale is die tentakels bedags teruggetrek en snags uitgesprei om plankton en ander klein organismes te vang. Beide sagte en steenkorale in vlak water kan hul planktondieet aanvul met die produkte van fotosintese wat moontlik gemaak word deur die simbiose met eensellige alge.[10] Die poliepe is verbind met ’n komplekse en goed ontwikkelde stelsel gastrovaskulêre kanale, wat meebring dat voedingstowwe en simbionte maklik gedeel kan word.[12]
Ekologie
wysigPoliepe lewe van ’n verskeidenheid klein organismes, van mikroskopiese plankton tot klein vissies. Die poliep se tentakels verlam die prooi of maak dit dood deur middel van netelselle. Die selle bevat gif wat hulle vinnig vrystel wanneer ’n ander organisme daaraan raak – gifangels word in die prooi geskiet en die gif word deur die hol filament vrygestel. Die verlamde prooi word dan deur die tentakels in die mondopening geplaas.[13]
Die tentakels trek dan saam om die prooi na die maag te bring. Wanneer die prooi verteer is, gaan die maag weer oop en die reste van die prooi word uitgeskei.
Baie korale, sowel as ander lede van Cnidaria soos seeanemone, vorm ’n simbiotiese verhouding met ’n klas alge, soöxantellas, van die genus Symbiodinium.[14] Elke poliep huisves gewoonlik een spesie alge. Deur fotosintese verskaf die alge energie aan die korale en help met verkalking.[15] Tot 30% van die weefsel van ’n poliep kan uit alge bestaan.[14]:23
Die voordeel vir die alge is dat hulle ’n veilig blyplek het en die poliep se afval-koolstofdioksied en -stikstof inneem. Die spanning wat die alge op die poliepe kan plaas, word dikwels verlig deurdat die poliepe die alge uitwerp. Massa-uitwerpings staan bekend as koraalverbleiking, want die alge dra by tot die koraal se bruin kleur; ander kleure is egter vanweë gaskoraalpigmente, soos groen fluoresseerproteïene. Uitwerpings verhoog die poliep se kans om korttermynspanning te oorleef – hulle kan altyd later weer alge opneem, miskien van ’n ander spesie. As die spanningstoestande egter voortduur, kan die poliep eindelik doodgaan.[16]
Voortplanting
wysigKorale kan beide gonochoristies (eenslagtig) en hermafrodities (tweeslagtig) wees; elk kan óf seksueel óf aseksueel voortplant. Voortplanting stel korale in staat om hulle op nuwe plekke te vestig.
Seksueel
wysigDie meeste korale plant seksueel voort. Sowat 25% van tweeslagtige korale (steenkorale) vorm gonochoristiese (enkelsel)-kolonies, terwyl die res tweeslagtig is.[17]
Uitstrooiers
wysig
Sowat 75% van alle hermatipiese (rifbouende) korale strooi hul saad uit deur gamete – eiers en sperma – in die water vry te stel. Die gamete verbind en vorm ’n mikroskopiese larwe, ’n planula, wat gewoonlik pienk en ovaalvormig is. ’n Tipiese koraalkolonie produseer miljoene larwes per jaar om die kans te vergroot dat ’n nuwe kolonie vorm.[18]
Sinkroniese uitstrooiing is tipies van ’n koraalrif. Dikwels, al is verskeie spesies teenwoordig, strooi al die korale hul saad dieselfde nag uit. Dit is noodsaaklik sodat manlike en vroulike gamete by mekaar kan uitkom. Die tyd vir uitstrooiing hang van omgewingsfaktore af wat van spesie tot spesie verskil. Dit kan temperatuurveranderings, die maansiklus, daglengte of chemiese tekens wees.[17] Sinkroniese uitstrooiing kan tot verbastering lei en is moontlik betrokke by koraalspesievorming.[19] Die geleentheid kan visueel dramaties wees, met die gewoonlik helder water wat troebel word van die gamete.
Broeiers
wysigSpesies wat broei, is gewoonlik ahermatipies (nierifvormend) in gebiede met sterk strome of groot branders. Hulle stel net sperma vry en dit sak af op die wagtende draers van onbevrugte eiers. Sinkroniese vrystelling van sperma kan ook by dié spesies plaasvind.[17] Ná bevrugting stel die korale planulas vry wat gereed is om by die kolonie aan te sluit.[15]
Planulas
wysigPlanulas (plat, swemmende larwes) swem na die lig om by die oppervlak van die water te kom, waar hulle dryf en groei voordat hulle afsak en ’n harde oppervlak soek waaraan hulle kan vasklou om ’n nuwe kolonie te begin. Hulle kan ook swem na die klank wat van die koraalrif af kom, en dus weg van diep waters.[20] Ondanks die groot getal gamete wat vrygestel word, vorm min nuwe kolonies omdat baie faktore daarteen tel. Die tyd van vrystelling tot afsakking is gewoonlik twee tot drie dae, maar kan tot twee maande wees.[21] Die larwes groei tot poliepe en word eindelik ’n koraalhoof deur aseksuele uitloping en groei.
Aseksueel
wysig
In ’n koraalhoof plant die geneties identiese poliepe aseksueel voort, óf deur uitloping (knopvorming) óf deur verdeling (in die lengte of breedte).
By uitloping ontstaan ’n jong poliep uit ’n volwasse een.[22] Terwyl die nuwe poliep groei, vorm sy liggaamsdele. Die afstand tussen die nuwe en ou poliep word groter en daarmee saam die senokarp (die weefsel wat die poliepe bind).
By verdeling ontstaan twee poliepe wat elk so groot word soos die oorspronklike een. As dit in die lengte verdeel, word die poliep breër en verdeel sy liggaam. Die mond verdeel ook en nuwe tentakels vorm. Die twee nuwe poliepe skep dan die liggaamsdele en eksoskelet wat hulle kort. As dit in die breedte gebeur, het een die voetgedeelte en die ander een die mondgedeelte; hulle groei dan albei die dele wat hulle kort.
Kolonieverdeling
wysigHele kolonies kan aseksueel voortplant deur twee kolonies met dieselfde genotipe te vorm. Die moontlike metodes sluit in splitsing, verlating en fragmentasie. Splitsing kom voor by sommige korale, veral in die familie Fungiidae, wanneer die kolonie in die vroeë ontwikkelingstadiums in twee of meer nuwe kolonies split. Verlating gebeur wanneer ’n enkele poliep die kolonie verlaat en hom aan ’n ander substrata anker om ’n nuwe kolonie te vorm. Fragmentasie vind plaas wanneer individuele poliepe tydens ’n storm of ander ontwrigting van die kolonie afbreek. Hulle kan dan ’n nuwe kolonie begin.[23]
Koraalriwwe
wysig- Die hoofartikel vir hierdie afdeling is: Koraalrif.

Baie korale in die orde Scleractinia is hermatipies, wat beteken hulle bou riwwe. Die meeste van dié korale verkry van hul energie van alge van die genus Symbiodinium bekend as soöxantellas. Hulle leef in ’n simbiotiese verhouding met hierdie fotosintetiese alga wat sonlig opneem; daarom word rifbouende korale meestal in vlak water aangetref. Hulle skei kalsiumkarbonaat af en vorm harde skelette wat die raamwerk van die rif word.

Nie alle rifbouende korale in vlak water bevat egter soöxantellas nie, en sommige diepwaterspesies wat voorkom op plekke waar daar nie sonlig kom nie, bou riwwe maar bevat nie soöxantellas nie.[24]
Daar is verskeie soorte vlakwaterkoraalriwwe, onder meer randriwwe, sperriwwe en atolle; die meeste kom in tropiese en subtropiese waters voor. Hulle groei besonder stadig, teen miskien ’n sentimeter per jaar in hoogte. Die Groot Koraalrif by Australië het vermoedelik twee miljoen jaar gelede begin vorm. Mettertyd verdeel koraalriwwe en gaan korale dood, sand en afval versamel tussen die korale en die skulpe van mossels en ander weekdiere verrot en vorm ’n geleidelik ontwikkelende struktuur van kalsiumkarbonaat.[25]
Koraalriwwe is uiters diverse mariene ekostelsels wat meer as 4 000 spesies visse huisves, asook enorme hoeveelhede netel-, week-, skaal- en ander soorte diere.[26]
Evolusiegeskiedenis
wysigHoewel korale die eerste keer in die Kambrium-periode, sowat 542 miljoen jaar gelede, voorgekom het,[27] is fossiele uiters skaars tot met die Ordovisium-periode, 100 miljoen jaar later, toe korale van die (nou uitgestorwe) ordes Rugosa en Tabulata algemeen geword het. Paleosoïese korale het dikwels talle simbionte in hul weefsel bevat.[28][29]
Tabulata-korale het in kalksteen en kalkagtige leiaarde van die periodes Ordovisium en Siluur voorgekom. Hul getalle het begin afneem in die middel van die Siluur en hulle het aan die einde van die Perm-periode, sowat 250 miljoen jaar gelede, uitgesterf.[30]
Rugosa-korale het in die middel van die Siluur begin oorheers en vroeg in die Trias-periode uitgesterf. Hulle het alleen en in kolonies voorgekom en het ook uit kalsiet bestaan.[31]
Die steenkorale het die nis gevul wat deur die uitgestorwe spesies Rugosa en Tabulata oopgelaat is. Hul fossiele kan in klein getalle in rotse van die Trias gevind word en het in die Jura en latere periodes algemeen geraak.[32] Hoewel hulle geologies jonger as die Rugosa en Tabulata is, word die aragoniet in hul skelette moeiliker gepreserveer, en hul fossielrekord is dus minder volledig.
 | |
|
Tydlyn van die grootste koraalfossielrekords en -ontwikkelings van 650 m.j.g. (miljoen jaar gelede) tot nou.[33][34] | |
Op sekere tye in die geologiese verlede was korale baie volop. Nes moderne korale het hul voorsate riwwe geskep, waarvan baie groot strukture in afsettingsgesteentes vorm. Saam met hulle is daar ook fossiele van ander rifdiere soos alge en sponse. Dit maak sommige korale nuttige gidsfossiele.[35]
Koraalfossiele is nie beperk tot rifoorblyfsels nie, en baie fossiele wat alleen voorkom, word elders ontdek.
Status
wysigBedreiging
wysig
Koraalriwwe word wêreldwyd bedreig.[36] Koraalwinning, landbou- en stedelike afvalprodukte, besoedeling (organies en anorganies), oorbevissing, siektes en die grawe van kanale om toegang tot eilande en baaie te kry stel die voortbestaan van koraalriwwe in gevaar. Ander bedreigings sluit in die styging van seetemperature en -watervlakke asook pH-veranderings weens suurvorming in oseane, faktore wat alles met die afskeiding van kweekhuisgasse verband hou.[37]
Sowat 10% van die wêreld se koraalriwwe is reeds dood[38][39][40] en sowat 60% word deur menslike bedrywighede bedreig.[41] Riwwe is veral in gevaar in Suidoos-Asië, waar 80% bedreig word.[42] Meer as 50% van die wêreld se koraalriwwe kan teen 2030 vernietig wees; daarom beskerm die meeste lande dit deur middel van omgewingswette.[43]
In die Karibiese See en tropiese Stille Oseaan veroorsaak direkte kontak tussen korale en seewiere bleiking en die dood van die koraal deur die oordrag van lipiedoplosbare metaboliete (stofwisselingsprodukte).[44] Seewier en alge floreer wanneer hulle genoeg voedingstowwe kry en daar nie baie plantetende roofdiere soos papegaaivisse in die omgewing is nie.
’n Verandering van meer as 1-2 °C in die watertemperatuur en ’n verhoging van die soutgehalte kan sekere soorte koraal laat doodgaan. Onder sulke spanning werp die korale hul soöxantellas uit en daarsonder toon die koraalweefsel die wit van hul skelette – dit staan as koraalbleiking bekend.[45]
Ondersese fonteine aan die kus van die Yucatán-skiereiland in Mexiko produseer water met ’n natuurlike lae pH (’n redelik hoë suurgehalte) en dit skep toestande wat ooreenstem met dié wat na verwagting algemeen sal word namate oseane koolstofdioksied absorbeer.[46] In opnames is verskeie spesies koraal gevind wat oënskynlik die suurgehalte verdra. Die kolonies was egter klein en verspreid.[46]
Beskerming
wysigMariene bewaringsgebiede, biosfeerreservate, seeparke, wêrelderfenisgebiede, visserybestuur en habitatbeskerming kan koraalriwwe teen besoedelingskade beskerm.[47]
Baie regerings verbied nou die verwydering van koraal van riwwe en lig kusbewoners in oor rifbeskerming en ekologie. Hoewel plaaslike bedrywighede soos habitatherstel en herbivoorbeskerming plaaslike skade kan verminder, bly langtermynbedreigings soos versuring en temperatuur- en seevlakstygings ’n probleem.[37]
Om die vernietiging van koraalriwwe in hul inheemse habitats te verminder, is projekte begin om koraal in nietropiese lande te groei.[48][49]
Die mens se rol
wysigPlaaslike ekonomieë naby groot koraalriwwe trek voordeel uit die groot hoeveelhede vis en ander seediere as ’n voedselbron. Riwwe skep ook geleenthede vir skubaduik- en snorkel-toerisme. Dié bedrywighede kan koraal beskadig, maar internasionale projekte soos Green Fins wat duik- en snorkelsentrums aanmoedig om ’n gedragskode te aanvaar, het al ’n positiewe uitwerking gehad.[50]
Lewende korale is baie gesog vir akwariums. Sagte korale is makliker om in aanhouding te versorg as harde korale.[51]
Juweliersware
wysig,_ca._1920s,_71.57.1.jpg)
Korale se helder kleure maak hulle gewilde items vir halssnoere en ander juweliersware. Intense rooi koraal word beskou as ’n edelsteen. Dit is baie skaars omdat dit al te veel geoes is.[52]
Die Chinese het rooi koraal nog altyd as ’n waardevolle mineraal beskou en dit verbind met goeie vooruitsigte en ’n lang lewe.[53] Die gewildheid daarvan het ’n hoogtepunt bereik tydens die Qing-dinastie (1644–1911), toe dit feitlik uitsluitlik vir juwele en miniatuur-mineraalboompies vir die keiser gebruik is. Die vroeë moderne handel in koraal, wat in Chinees as shanhu bekend is, het in die Middellandse See begin en is deur die Britse Oos-Indiese Kompanjie na Sjina uitgebrei.[54]
Medisyne
wysig
In geneeskunde word chemiese samestellings van korale gebruik vir die behandeling van kanker, vigs en pyn, asook vir ander doeleindes. Koraalskelette, byvoorbeeld Isididae, word ook gebruik vir beenoorplantings in mense.[56]
Coral Calx, bekend as Praval Bhasma in Sanskrit, word algemeen in tradisionele Indiese medisyne gebruik as ’n aanvulling in die behandeling van verskeie beenmetabolisme-afwykings wat met ’n kalsiumtekort verband hou.[57] In klassieke tye is die inneem van fyngemaalde koraal, wat hoofsaaklik uit kalsiumkarbonaat bestaan het, aanbeveel vir die behandeling van maagsere.[58]
Boumateriaal
wysigKoraalriwwe in plekke soos die kus van Oos-Afrika word gebruik as ’n bron van boumateriaal.[59] Antieke fossiele koraalkalksteen, soos dié van die heuwels om Oxford in Engeland, is eens gebruik as boustene en kan in ’n paar van die oudste geboue in die stad gesien word, asook in die Middeleeuse stadsmure.[60]
Klimaatnavorsing
wysigJaarlikse groeiringe in sommige korale soos die diepsee-bamboeskoraal Isididae, kan van die eerste tekens toon van die uitwerking van oseaanversuring op die seelewe.[61] Die groeiringe stel geoloë in staat om jaar-vir-jaar-opnames, ’n soort datering, te doen wat as grondslag dien van hoëresolusierekords van klimaats- en omgewingsveranderings van die verlede met die hulp van geochemiese tegnieke.[62]
Sekere spesies vorm kolonies wat mikro-atols genoem word: kolonies wat bo dood is en bo die watervlak uitsteek, maar waarvan die kante meestal onderwater en lewend is. Gemiddelde getyvlakke beperk hul hoogte. Deur verskillende groeimorfologieë te bestudeer, kan ’n laeresolusierekord van seevlakveranderings opgestel word. Die ouderdom van fossiele mikro-atols kan ook met behulp van radiokoolstofdatering vasgestel word. Sulke metodes kan help om Holoseen-seevlakke te rekonstrueer.[63]
Toenemende seetemperature in tropiese streke (~1 ºC) in die afgelope eeu het koraalbleiking, die dood en dus die krimping van koraalbevolkings meegebring. Hoewel korale kan aanpas en akklimatiseer, is dit onseker of hierdie evolusieproses vinnig genoeg sal plaasvind om ’n groot afname in hul getalle te voorkom.[64]
Hoewel koraal groot bevolkings het wat seksueel voortpant, kan hul evolusie verlangsaam word deur oorvloedige aseksuele voortplanting.[65] Geenvloei (die uitruil van allele en gene tussen bevolkings) verskil van spesie tot spesie.[65] Volgens die biogeografie van koraalspesies kan nie op geenvloei staatgemaak word as ’n betroubare bron van aanpassing nie omdat hulle baie onbeweeglike organismes is. Die lengte van korale se lewe kan ook hul aanpassing beïvloed.[65]
Aanpassing by klimaatsveranderings is egter al in baie gevalle geopenbaar. Dit vind gewoonlik plaas vanweë ’n verandering in koraal- en soöxantella-genotipes. Hierdie veranderings in alleelfrekwensies het gelei tot meer verdraagsame tipes soöxantellas.[66] Wetenskaplikes het bevind ’n sekere soort steenkoraal-soöxantella word meer algemeen waar die seetemperatuur hoog is.[67][68]
In die Golf van Mexiko, waar seetemperature styg, het kouesensitiewe horingkorale en hul simbionte nie net geskuif nie,[66] maar dit lyk of daar ’n sekere groeitempo is wat gunstig is vir seleksie. Stadiger groeiende korale wat meer hittebestand is, het toegeneem.[69]
Akwariums
wysig
Soutwaterakwariums het die afgelope tyd baie uitgebrei en sluit nou vistenks in met groot hoeveelhede "lewende rotse" waarop korale toegelaat word om te groei en versprei.[70] ("Lewende rotse" verwys na klippe uit die see wat bestaan uit skelette van dooie organismes.) Hierdie tenks word óf in ’n natuurlike toestand gehou met alge en ’n diep sandbed wat filtrasie verskaf[71] óf as "vertoontenks", met die rotse wat skoongehou word van die alge en mikrofauna wat normaalweg daarop sou groei[72] – dit word gedoen sodat die tenk skoon en netjies kan bly.

Sagte koraal is die gewildste vir rifakwariums, want van hulle is maklik om te groei en plant voort in ’n verskeidenheid toestande.[73] Ernstiger visliefhebbers hou soms steenkoraal met klein poliepe in hul tenks, al is dit moeilik om te versorg. Steenkoraal met groot poliepe is ’n goue middeweg.
Koraalboerdery
wysigKoraalboerdery is die groei van korale vir onder meer navorsings- en handelsdoeleindes of vir die herstel van koraalriwwe. Veral vir laasgenoemde toon koraalboerdery groot potensiaal, omdat riwwe wêreldwyd bedreig word.[74][75][76]
Die proses behels die verkryging van stukkies koraal of larwes van ’n koraalrif. Dit is algemeen bekend as die "tuinmaakmetode" omdat dit ooreenstem met die groei van plante in ’n kwekery totdat hulle groot genoeg is om uitgeplant te word.[76][77] Die koraal word dus eintlik soos ’n plant behandel en die dele wat in koraalkwekerye gegroei word, word dan ook "sade" genoem. Hulle word daarna in hul natuurlike habitat ontplooi, in dié geval die see.[76] So word die vroeë natuurlike groeistadiums van korale, wanneer hulle die meeste geneig is om dood te gaan, gesystap.
Koraalboerdery word hoofsaaklik om drie redes beoefen: bewaring, om openbare akwariums te verskaf vir uitstallings en om materiaal aan privaat akwariums te verskaf. Volgroeide korale kan ook teruggeplaas word op ’n koraalrif, gewoonlik waar dit beskadig is, om die herstel van die rif aan te help.[76][78]
Galery
wysig-
 Skelet van ’n Fungia-spesie.
Skelet van ’n Fungia-spesie. -
 Poliepe van Eusmilia fastigiata.
Poliepe van Eusmilia fastigiata. -
 Dendrogyra cylindricus.
Dendrogyra cylindricus. -
 Diploria labyrinthiformis.
Diploria labyrinthiformis. -
 Koraal stel saad vry.
Koraal stel saad vry. -
 Koraal stel eiers vry.
Koraal stel eiers vry. -
 ’n Randkoraalrif aan die kus van Israel.
’n Randkoraalrif aan die kus van Israel.
_Top_Macro_91.JPG)

_(San_Salvador_Island,_Bahamas)_1_(15513345363).jpg)




Verwysings
wysig- ↑ Hoeksema, Bert (2015). "Octocorallia". World Register of Marine Species. Besoek op 24 April 2015.
- ↑ Hoeksema, Bert (2015). "Hexacorallia". World Register of Marine Species. Besoek op 24 April 2015.
- ↑ Hoeksema, Bert (2016). "Ceriantharia". World Register of Marine Species. Besoek op 4 Februarie 2017.
- ↑ Squires, D.F. (1959). "Deep sea corals collected by the Lamont Geological Observatory. 1. Atlantic corals". American Museum Novitates. 1965: 1–42.
- ↑ Hoeksema, Bert (2015). "Anthozoa". World Register of Marine Species. Besoek op 24 April 2015.
- ↑ Ceriantharia, deur Hoeksema, Bert. WoRMS, 2016. URL besoek op 18 Mei 2017
- ↑ Stampar, S.N.; Maronna, M.M.; Kitahara, M.V.; Reimer, J.D.; Morandini, A.C. (2014). "Fast-Evolving Mitochondrial DNA in Ceriantharia: A Reflection of Hexacorallia Paraphyly?". PLoS ONE. 9 (1): e86612. doi:10.1371/journal.pone.0086612. PMC 3903554. PMID 24475157.
- ↑ Chen, C. A.; Odorico, D. M.; ten Lohuis, M.; Veron, J. E. N.; Miller, D. J. (1995). "Systematic relationships within the Anthozoa (Cnidaria: Anthozoa) using the 5'-end of the 28S rDNA" (PDF). Molecular Phylogenetics and Evolution. 4 (2): 175–183. doi:10.1006/mpev.1995.1017. PMID 7663762. Geargiveer vanaf die oorspronklike (PDF) op 2 April 2008. Besoek op 24 Augustus 2021.
- ↑ Milleporidae Fleming, 1828, deur Schuchert, Peter. WoRMS, 2015. URL besoek op 18 Mei 2017
- ↑ 10,0 10,1 10,2 Ruppert, Edward E.; Fox, Richard, S.; Barnes, Robert D. (2004). Invertebrate Zoology, 7de uitg. Cengage Learning. pp. 132–148. ISBN 978-81-315-0104-7.
{{cite book}}: AS1-onderhoud: meer as een naam (link) - ↑ Sprung, Julian (1999). Corals: A quick reference guide. Ricordea Publishing. p. 145. ISBN 1-883693-09-8.
- ↑ D. Gateno; A. Israel; Y. Barki; B. Rinkevich (1998). "Gastrovascular Circulation in an Octocoral: Evidence of Significant Transport of Coral and Symbiont Cells". The Biological Bulletin. Marine Biological Laboratory. 194 (2): 178–186. doi:10.2307/1543048. JSTOR 1543048.
- ↑ "Coral Feeding Habits" (in Engels). NOAA. Geargiveer vanaf die oorspronklike op 9 Oktober 2019. Besoek op 25 April 2015.
- ↑ 14,0 14,1 Murphy, Richard C. (2002). Coral Reefs: Cities Under The Seas. The Darwin Press. ISBN 0-87850-138-X.
- ↑ 15,0 15,1 Madl, P.; Yip, M. (2000). "Field Excursion to Milne Bay Province – Papua New Guinea" (in Engels). Geargiveer vanaf die oorspronklike op 11 Mei 2020. Besoek op 31 Maart 2006.
- ↑ W. W. Toller; R. Rowan; N. Knowlton (2001). "Repopulation of Zooxanthellae in the Caribbean Corals Montastraea annularis and M. faveolata following Experimental and Disease-Associated Bleaching". The Biological Bulletin. Marine Biological Laboratory. 201 (3): 360–373. doi:10.2307/1543614. JSTOR 1543614. PMID 11751248.
- ↑ 17,0 17,1 17,2 Veron, J.E.N. (2000). Corals of the World. Vol 3 (3rd uitg.). Australia: Australian Institute of Marine Sciences and CRR Qld Pty Ltd. ISBN 0-642-32236-8.
- ↑ How Do Corals Reproduce?, NOAA Ocean Service Education
- ↑ Hatta, M.; Fukami, H.; Wang, W.; Omori, M.; Shimoike, K.; Hayashibara, T.; Ina, Y.; Sugiyama, T. (1999). "Reproductive and genetic evidence for a reticulate evolutionary theory of mass spawning corals" (PDF). Molecular Biology and Evolution. 16 (11): 1607–1613. doi:10.1093/oxfordjournals.molbev.a026073. PMID 10555292.
- ↑ Vermeij, Mark J. A.; Marhaver, Kristen L.; Huijbers, Chantal M.; Nagelkerken, Ivan; Simpson, Stephen D. (2010). "Coral Larvae Move toward Reef Sounds". PLoS ONE. 5 (5): e10660. doi:10.1371/journal.pone.0010660. PMC 2871043. PMID 20498831.
{{cite journal}}: Onbekende parameter|laydate=geïgnoreer (hulp); Onbekende parameter|laysource=geïgnoreer (hulp); Onbekende parameter|laysummary=geïgnoreer (hulp) - ↑ Jones, O.A.; R. Endean. (1973). Biology and Geology of Coral Reefs. New York, USA: Harcourt Brace Jovanovich. pp. 205–245. ISBN 0-12-389602-9.
- ↑ Barnes, R. and; Hughes, R. (1999). An Introduction to Marine Ecology (3rd uitg.). Malden, MA: Blackwell. pp. 117–141. ISBN 0-86542-834-4.
- ↑ Sheppard, Charles R.C.; Davy, Simon K.; Pilling, Graham M. (25 Junie 2009). The Biology of Coral Reefs. OUP Oxford. pp. 78–81. ISBN 978-0-19-105734-2.
- ↑ Schuhmacher, Helmut; Zibrowius, Helmut (1985). "What is hermatypic?". Coral Reefs. 4 (1): 1–9. Bibcode:1985CorRe...4....1S. doi:10.1007/BF00302198.
- ↑ MSN Encarta (2006). "Great Barrier Reef".. URL besoek op 25 April 2015. Geargiveer 28 Oktober 2009 op Wayback Machine
- ↑ Spalding, Mark; Ravilious, Corinna; Green, Edmund (2001). World Atlas of Coral Reefs. Berkeley, CA: University of California Press and UNEP/WCMC. pp. 205–245. ISBN 0-520-23255-0.
- ↑ Pratt, B.R.; Spincer, B.R., R.A. Wood and A.Yu. Zhuravlev (2001). "12: Ecology and Evolution of Cambrian Reefs". Ecology of the Cambrian Radiation (PDF). Columbia University Press. p. 259. ISBN 0-231-10613-0. Besoek op 6 April 2007.
{{cite book}}: AS1-onderhoud: meer as een naam (link) - ↑ Vinn, O.; Mõtus, M.-A. (2008). "The earliest endosymbiotic mineralized tubeworms from the Silurian of Podolia, Ukraine". Journal of Paleontology. 82 (2): 409–414. doi:10.1666/07-056.1. Besoek op 11 Junie 2014.
- ↑ Vinn, O.; Mõtus, M.-A. (2012). "Diverse early endobiotic coral symbiont assemblage from the Katian (Late Ordovician) of Baltica". Palaeogeography, Palaeoclimatology, Palaeoecology. 321–322: 137–141. doi:10.1016/j.palaeo.2012.01.028. Besoek op 11 Junie 2014.
- ↑ "Introduction to the Tabulata" (in Engels). UCMP Berkeley. Geargiveer vanaf die oorspronklike op 8 Mei 2020. Besoek op 25 April 2015.
- ↑ "Introduction to the Rugosa" (in Engels). UCMP Berkeley. Geargiveer vanaf die oorspronklike op 9 Mei 2020. Besoek op 25 April 2015.
- ↑ "Evolutionary history". AIMS. Besoek op 25 April 2015.
- ↑ Ben M. Waggoner (2000). "Anthozoa: Fossil Record". Anthozoa. UCMP. http://www.ucmp.berkeley.edu/cnidaria/anthozoafr.html. Besoek op 2009-03-23.
- ↑ William A. Oliver, Jr. (2003). "Corals: Table 1". Fossil Groups. USGS. http://geology.er.usgs.gov/paleo/corals.shtml. Besoek op 2009-03-23.
- ↑ Alden, Andrew. "Index Fossils" (in Engels). About education. Geargiveer vanaf die oorspronklike op 1 Desember 2016. Besoek op 25 April 2015.
- ↑ "Coral reefs around the world" (in Engels). Guardian.co.uk. 2 September 2009. Geargiveer vanaf die oorspronklike op 18 April 2020.
- ↑ 37,0 37,1 "Threats to Coral Reefs". Coral Reef Alliance. 2010. Geargiveer vanaf die oorspronklike op 1 Desember 2011. Besoek op 5 Desember 2011.
- ↑ Kleypas, J.A.; Feely, R.A.; Fabry, V.J.; Langdon, C.; Sabine, C.L.; Robbins, L.L. (2006). "Impacts of Ocean Acidification on Coral Reefs and Other Marine Calcifiers: A guide for Future Research" (PDF). National Science Foundation, NOAA, & United States Geological Survey. Geargiveer vanaf die oorspronklike (PDF) op 20 Julie 2011. Besoek op 7 April 2011.
{{cite journal}}: Cite journal requires|journal=(hulp) - ↑ Save Our Seas, 1997 Summer Newsletter, drr. Cindy Hunter en Alan Friedlander
- ↑ Tun, K.; Chou, L.M.; Cabanban, A.; Tuan, V.S.; Philreefs; Yeemin, T.; Suharsono; Sour, K.; Lane, D. (2004). "Status of Coral Reefs, Coral Reef Monitoring and Management in Southeast Asia, 2004". In Wilkinson, C. (red.). Status of Coral Reefs of the world: 2004. Townsville, Queensland, Australië: Australian Institute of Marine Science. pp. 235–276.
- ↑ Burke, Lauretta; Reytar, K.; Spalding, M.; Perry, A. (2011). Reefs at risk revisited. Washington, DC: World Resources Institute. p. 38. ISBN 978-1-56973-762-0.
- ↑ Bryant, Dirk; Burke, Lauretta; McManus, John; Spalding, Mark. "Reefs at Risk: A Map-Based Indicator of Threats to the World's Coral Reef" (PDF). NOAA. Geargiveer vanaf die oorspronklike (PDF) op 18 Februarie 2013. Besoek op 25 April 2015.
- ↑ Norlander (8 Desember 2003). "Coral crisis! Humans are killing off these bustling underwater cities. Can coral reefs be saved? (Life science: corals)". Science World.
- ↑ Rasher DB, Hay ME; Hay (Mei 2010). "Chemically rich seaweeds poison corals when not controlled by herbivores". Proceedings of the National Academy of Sciences of the United States of America. 107 (21): 9683–8. doi:10.1073/pnas.0912095107. PMC 2906836. PMID 20457927.
- ↑ Hoegh-Guldberg, O. (1999). "Climate change, coral bleaching and the future of the world's coral reefs" (PDF). Marine and Freshwater Research. 50 (8): 839–866. doi:10.1071/MF99078. Geargiveer vanaf die oorspronklike (PDF) op 26 April 2012. Besoek op 20 Mei 2017.
- ↑ 46,0 46,1 Stephens, Tim (28 November 2011). "Submarine springs offer preview of ocean acidification effects on coral reefs" (in Engels). University of California Santa Cruz. Geargiveer vanaf die oorspronklike op 10 Mei 2020. Besoek op 25 April 2015.
- ↑ "Phoenix Rising". National Geographic Magazine. Januarie 2011. Besoek op 30 April 2011.
- ↑ EcoDeco EcologicalTechnology Geargiveer 7 Maart 2011 op Wayback Machine. Ecodeco.nl. URL besoek op 2011-11-29.
- ↑ KoralenKAS project Geargiveer 26 April 2012 op Wayback Machine. Koraalwetenschap.nl. URL besoek op 29 November 2011.
- ↑ Hunt, Chloe V.; Harvey, James J.; Miller, Anne; Johnson, Vivienne; Phongsuwan, Niphon (2013). "The Green Fins approach for monitoring and promoting environmentally sustainable scuba diving operations in South East Asia". Ocean & Coastal Management. 78: 35–44. doi:10.1016/j.ocecoaman.2013.03.004.
- ↑ "Eight great soft corals for new reefkeepers" (in Engels). AquaDaily. 5 Desember 2008. Geargiveer vanaf die oorspronklike op 2 Desember 2018. Besoek op 2 Januarie 2009.
- ↑ Magsaysay, Melissa (21 Junie 2009). "Coral makes a splash". Los Angeles Times. Besoek op 12 Januarie 2013.
- ↑ Welch, Patricia Bjaaland, Chinese Art: A Guide to Motifs and Visual Imagery. Tokyo, Rutland and Singapore: Tuttle, 2008, bl. 61
- ↑ Lacey, Pippa, "The Coral Network: The trade of red coral to the Qing imperial court in the eighteenth century" in The Global Lives of Things, reds. Anne Gerritsen en Giorgio Aiello, Londen: Rutledge, 2016, bl. 81
- ↑ Folio 391, Juliana Anicia Codex
- ↑ H. Ehrlich, P. Etnoyer, S. D. Litvinov; Etnoyer, P.; Litvinov, S. D.; Olennikova, M.M.; Domaschke, H.; Hanke, T.; Born, R.; Meissner, H.; Worch, H.; et al. (2006). "Biomaterial structure in deep-sea bamboo coral (Anthozoa: Gorgonacea: Isididae)". Materialwissenschaft und Werkstofftechnik. www3.interscience.wiley.com. 37 (6): 552–557. doi:10.1002/mawe.200600036. Besoek op 11 Mei 2009.
{{cite journal}}: AS1-onderhoud: meer as een naam (link)[dooie skakel] - ↑ Reddy PN, Lakshmana M, Udupa UV; Lakshmana; Udupa (Desember 2003). "Effect of Praval bhasma (Coral calx), a natural source of rich calcium on bone mineralization in rats". Pharmacological Research. 48 (6): 593–9. doi:10.1016/S1043-6618(03)00224-X. PMID 14527824.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Pedanius Dioscorides – Der Wiener Dioskurides, Codex medicus Graecus 1 der Österreichischen Nationalbibliothek Graz: Akademische Druck- und Verlagsanstalt 1998 fol. 391 verso (Band 2), Kommentar S. 47 und 52. ISBN 3-201-01725-6
- ↑ Pouwels, Randall L. (6 Junie 2002). Horn and Crescent: Cultural Change and Traditional Islam on the East African Coast, 800–1900. Cambridge University Press. p. 26. ISBN 978-0-521-52309-7.
- ↑ "Strategic Stone Study: A Building Stone Atlas of Oxfordshire". English Heritage. Maart 2011. Geargiveer vanaf die oorspronklike op 9 Oktober 2019. Besoek op 23 April 2015.
- ↑ "National Oceanic and Atmospheric Administration – New Deep-Sea Coral Discovered on NOAA-Supported Mission". www.noaanews.noaa.gov. Besoek op 11 Mei 2009.
- ↑ Schrag, D.P.; Linsley, B.K. (2002). "Corals, chemistry, and climate". Science. 296 (8): 277–278. doi:10.1126/science.1071561. PMID 11951026.
- ↑ Smithers, Scott G.; Woodroffe, Colin D. (2000). "Microatolls as sea-level indicators on a mid-ocean atoll". Marine Geology. 168 (1–4): 61–78. doi:10.1016/S0025-3227(00)00043-8.
- ↑ Hoegh-Guldberg O. (1999). "Climate change, coral bleaching and the future of the world's coral reefs". Marine and Freshwater Research. 50 (8): 839–99. doi:10.1071/mf99078.
- ↑ 65,0 65,1 65,2 Hughes, T.; Baird, A.; Bellwood, D.; Card, M.; Connolly, S.; Folke, C.; Grosberg, R.; Hoegh-Guldberg, O.; Jackson, J.; Klepas, J.; Lough, J.; Marshall, P.; Nystrom, M.; Palumbi, S.; Pandolfi, J.; Rosen, B.; and Roughgarden, J. (2003). "Climate change, human impacts, and the resilience of coral reefs". Science. 301 (5635): 929–33. doi:10.1126/science.1085046. PMID 12920289.
- ↑ 66,0 66,1 Parmesan, C. (2006). "Ecological and evolutionary responses to recent climate change". Annual Review of Ecology, Evolution, and Systematics. 37: 637–69. doi:10.1146/annurev.ecolsys.37.091305.110100.
- ↑ Baker, A. (2004). "Corals' adaptive response to climate change". Nature. 430 (7001): 741. doi:10.1038/430741a.
- ↑ Donner, S., Skirving, W., Little, C., Oppenheimer, M., and Hoegh-Guldenberg (2005). "Global assessment of coral bleaching and required rates of adaptation under climate change". Global Change Biology. 11 (12): 2251–65. doi:10.1111/j.1365-2486.2005.01073.x.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Baskett, M.; Gaines, S.; Nisbet, R. (2009). "Symbiont diversity may help coral reefs survive moderate climate change". Ecological Applications. 19 (1): 3–17. doi:10.1890/08-0139.1. PMID 19323170.
{{cite journal}}: Onbekende parameter|last-author-amp=geïgnoreer (hulp) - ↑ Aquarium Corals: Collection and Aquarium Husbandry of Northeast Pacific Non-Photosynthetic Cnidaria Geargiveer 6 Junie 2017 op Wayback Machine. Advancedaquarist.com (14 Januarie 2011). URL besoek op 13 Junie 2016.
- ↑ Reefkeeping 101 – Various Nutrient Control Methods. Reefkeeping.com. URL besoek op 2016-06-13.
- ↑ Aquarium Substrate & Live Rock Clean Up Tips. Saltaquarium.about.com. URL besoek op 13 Junie 2016.
- ↑ Coral Reefs. Marinebio.org. URL besoek op 13 Junie 2016.
- ↑ Horoszowski-Fridman YB, Izhaki I, Rinkevich B; Izhaki; Rinkevich (2011). "Engineering of coral reef larval supply through transplantation of nursery-farmed gravid colonies". Journal of Experimental Marine Biology and Ecology. 399 (2): 162–166. doi:10.1016/j.jembe.2011.01.005.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Pomeroy, Robert S.; Parks, John E.; Balboa, Cristina M. (2006). "Farming the reef: Is aquaculture a solution for reducing fishing pressure on coral reefs?". Marine Policy. 30 (2): 111–30. doi:10.1016/j.marpol.2004.09.001.
- ↑ 76,0 76,1 76,2 76,3 Rinkevich B (2008). "Management of coral reefs: We have gone wrong when neglecting active reef restoration" (PDF). Marine pollution bulletin. 56 (11): 1821–1824. doi:10.1016/j.marpolbul.2008.08.014. PMID 18829052. Geargiveer vanaf die oorspronklike (PDF) op 23 Mei 2013.
{{cite journal}}: Onbekende parameter|dead-url=geïgnoreer (hulp) - ↑ Levy, G, Shaish, L, Haim, A & Rinkevich, B (2010) "Mid-water rope nursery--Testing design and performance of a novel reef restoration instrument" Ecological Engineering, 36(4): 560–569.
- ↑ Bellwood DR et al. (2004) Confronting the coral reef crisis Nature, Review, 429(6994): 827–833
Eksterne skakels
wysig- Coral Reefs The Ocean Portal deur die Smithsonian-instituut
- NOAA CoRIS – Coral Reef Biology
- NOAA Ocean Service Education – Corals
- "Coral Factsheet". Waitt Institute. Geargiveer vanaf die oorspronklike op 3 Augustus 2016. Besoek op 4 Februarie 2017.
- "What is a coral?". Stanford microdocs project. Geargiveer vanaf die oorspronklike op 6 Januarie 2014. Besoek op 4 Februarie 2017.
 Wikispecies het meer inligting verwant aan Koraal
Wikispecies het meer inligting verwant aan Koraal Wikimedia Commons het meer media in die kategorie Koraal.
Wikimedia Commons het meer media in die kategorie Koraal.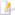 Hierdie artikel is vertaal uit die Engelse Wikipedia
Hierdie artikel is vertaal uit die Engelse Wikipedia